| 教員名 | 研究テーマ | メールアドレス ※アドレスの表記について |
|
| 岩口 伸一 | 菌類の染色体構造に関する分子生物学的研究 |  |
iwaguchi*cc.nara-wu.ac.jp |
| 鍵和田 聡 | 生体膜の構造と機能に関する分子生物学的研究 |  |
kagiwada*cc.nara-wu.ac.jp |
| 清水 隆之 | 光合成生物のレドックス応答と生理機能の調節に関する研究 | tshimizu*cc.nara-wu.ac.jp | |
| 杉浦 真由美 | 原生生物の生殖と細胞間相互作用に関する研究 |  |
msugi*cc.nara-wu.ac.jp |
| 西井 一郎 | 緑藻ボルボックス目の発生及び単細胞生物から多細胞生物への進化に関する分子生物学的研究 |  |
ichiron*cc.nara-wu.ac.jp |

| 教員名 | 研究テーマ | メールアドレス ※アドレスの表記について |
|
| 岡本 麻友美 | 脳組織づくりの原理を探る研究 |  |
mokamoto*cc.nara-wu.ac.jp |
| 川野 絵美 | 下等脊椎動物における非視覚系の光受容メカニズムに関する生理学的研究 |  |
kawano*cc.nara-wu.ac.jp |
| 酒井 敦 | 植物オルガネラの増殖・分化に関する生理・生化学的研究、植物個体の環境応答 |  |
sakai*cc.nara-wu.ac.jp |
| 高塚 大知 | 植物細胞の時空間ダイナミクス制御に関する研究・ バイオスティミュラントの開発 | 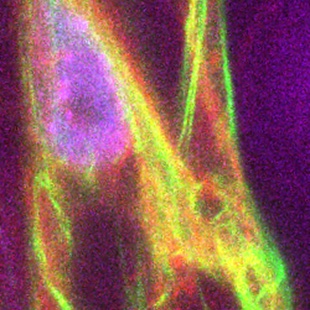 |
h-takatsuka*cc.nara-wu.ac.jp |
| 奈良 久美 | 植物の形態形成、及び水輸送に関する分子メカニズムの研究 |  |
kumisn*cc.nara-wu.ac.jp |
| 堀 沙耶香 | 行動最適化の原型回路モデルの動作原理に関する分子生理学的研究 |  |
hori*cc.nara-wu.ac.jp |
| 教員名 | 研究テーマ | メールアドレス ※アドレスの表記について |
|
| 井田 崇 | 植物の生理・繁殖生態および訪花昆虫の生態と行動に関する研究 |  |
tyida*cc.nara-wu.ac.jp |
| 上原 春香 | 鳥類等の脊椎動物における生態学的・心理学的行動研究 |  |
uehara*cc.nara-wu.ac.jp |
| 片野 泉 | 陸水生態系における生物多様性の維持メカニズムに関する研究 |  |
katano*cc.nara-wu.ac.jp |
| 京極 大助 | 有性生殖とその副次効果に関する進化生態学的な研究 |  |
|
| 遊佐 陽一 | 貝類を中心とする淡水・海洋生物の生態と行動の研究、およびそれにもとづく有害種の防除法の開発 |  |
yusa*cc.nara-wu.ac.jp |
| 1. 石油資化酵母の二形性変換(酵母⇔菌糸) 同一生物あるいは個体が2つの異なる形態をとることを一般に二形性 (dimorphism)と呼びます。菌類には生育環境により単細胞性の酵母型と多細胞性の菌糸型との増殖形態をとるものが知られ、この異なる形態を相互変換することから二形性変換 (dimorphic transition) と呼ばれています。不完全酵母Candida tropicalisは、土壌から分離される石油資化酵母で、生育環境を制御することにより、出芽による酵母型増殖と菌糸成長による菌糸型増殖の二つの異なる増殖形態をとります。この菌はグルコースを炭素源とした合成液体培地で培養すると、単細胞性の出芽増殖による酵母型を示しますが、グルコース合成培地にさらにエタノールを2.5%添加した培養では菌糸の成長が見られます。この酵母型から菌糸型への二形性変換のメカニズムを明らかにするために、細胞生物学的、分子生物学的手法を用いて研究を行っています。 
2. 植物(花)から分離される真菌(カビ、酵母)の研究  我たちの身の回りには非常に多くの真菌が生育していますが、食品や臨床的な面では研究が進んでいるのに比べ、野外環境に生育する真菌については知見が非常に乏しく、どういった場所にどういった種類の真菌が生息しているかについては詳しく分かっていません。環境真菌について理解するために、植物(花)から分離された真菌(カビ、酵母)について、DNAレベルの解析方法により同定し、研究を進めています。 我たちの身の回りには非常に多くの真菌が生育していますが、食品や臨床的な面では研究が進んでいるのに比べ、野外環境に生育する真菌については知見が非常に乏しく、どういった場所にどういった種類の真菌が生息しているかについては詳しく分かっていません。環境真菌について理解するために、植物(花)から分離された真菌(カビ、酵母)について、DNAレベルの解析方法により同定し、研究を進めています。(右図は、花酵母を分離するのに使われる天然記念物「奈良八重桜(ナラノヤエザクラ)」) |
| 生体膜は細胞や細胞小器官を他と区別するための障壁として働くばかりでなく、細胞の情報伝達・物質輸送・エネルギー生産の場として、またタンパク質などの生体分子の輸送・貯蔵・分解を行う特殊領域を作るために機能しています。 生体膜はおもにタンパク質と脂質から成りますが、これまで脂質分子については単なる構造を作るための要素と考えられていました。しかし最近では、脂質分子が特定の生理機能に関係することが明らかになってきており、それらの機能の解明が進んできています。 私たちは、生体膜の代表的な脂質であるリン脂質が、生体膜のどの部位でどのような生理現象に関係するか、リン脂質分子の局在や合成がどのように制御されているかを解明しようとしています。 生体膜のような多種類の分子集合体を、細胞から純粋に単離することは困難です。そこで、私たちは遺伝学的・分子生物学的な研究に適した出芽酵母(Saccharomyece cerevisiae)を用いて研究を行っています。 
|
|
【超硫黄分子に着目したレドックス応答と硫黄代謝】 「超硫黄分子」は、分子内に過剰な硫黄原子が付加された硫黄代謝物で、レドックス活性に富んだ分子です。 私たちは、硫黄要求性の紅色光合成細菌を用いて、超硫黄分子のセンシング機構とそれに関わる硫黄代謝系を明らかにすることで、超硫黄分子を考慮した真のレドックスシグナルに基づいた生命現象の理解を目指しています。 本研究は、レドックスバランスの破綻が引き起こす様々な疾病の新たな治療法開発につながる基本概念を創出する可能性を秘めています。 【植物における超硫黄分子シグナル伝達機構】 モデル植物シロイヌナズナを用いて、超硫黄分子が植物の生理機能におよぼす影響を調べることで、植物の成長やオルガネラ間情報伝達を、超硫黄分子シグナル制御機構を踏まえて再構築することを目指しています。 特に、光合成機能をつかさどる葉緑体の形成機構と超硫黄分子の関係に着目して研究を行っています。 また、農作物の生産力向上にむけた超硫黄分子のユニークな活用方法の開発も見据えて研究を行っています。 |
| 1.繊毛虫ブレファリズマにおける接合誘導機構と接合型決定機構に関する研究 ゾウリムシやブレファリズマなどの繊毛虫類は、栄養が豊富な環境下では二分裂を繰り返して増殖し、分裂回数を重ねるにつれて性的未熟期、成熟期、老衰期と発生段階を進行させる。栄養条件が悪化すると成熟期にある細胞は接合とよばれる有性生殖を開始する。接合は相補的な接合型細胞間で、交配フェロモンのような物質(ガモン)を介してお互いを刺激し合うことによって誘導される。接合を完了した細胞は新しい遺伝的組成をもった小核・大核をもち、それまでの分裂回数がリセットされ、次世代に相当する個体(細胞)となる。また、繊毛虫の接合型(性)の数は種によって様々であり、2種類のものから30種類以上のものまで知られている。繊毛虫において「どのような分子機構で接合が開始されるのか?」、また「接合型はどのように決まっているのか?」という疑問に答えるため、ブレファリズマを用いて主にガモンの構造や発現、機能に注目して研究を行っている。 2.ブレファリズマ属における交配フェロモンの多様性 繊毛虫類ブレファリズマ属には20種類の種の存在が知られているが、それらは主に大核の形態や体サイズなど形態の違いをもとに分類されたものである。そこで、これらの種の違いを遺伝子レベルで調べてブレファリズマの種の分類を見直すと共に、交配フェロモンであるガモンに注目し、異なる種間におけるガモンの分子構造や機能の比較を行い、ガモンの多様性と性的隔離、種の分化との関連を探ろうとしている。 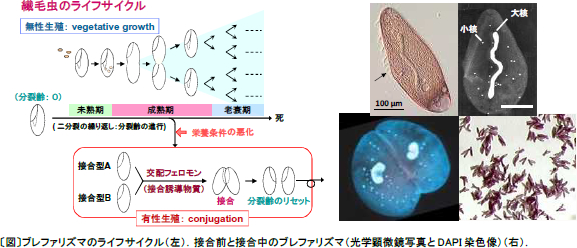 |
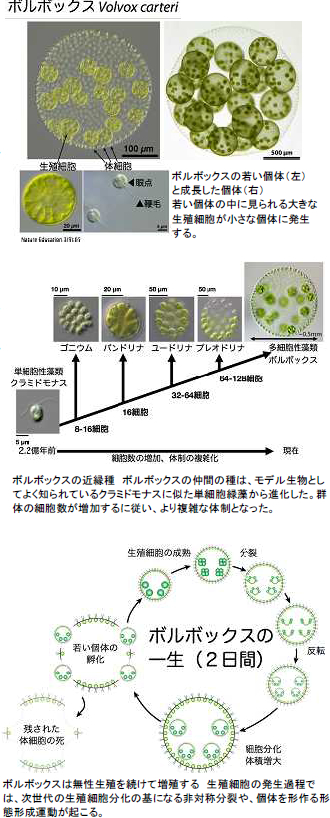 緑藻ボルボックス(直径0.1〜0.5 mm)は、球面を作る2千個の小さな体細胞と、約20個の大きな生殖細胞からなり、多細胞生物としては最もシンプルな体制をしている。ボルボックスの近縁種には、単細胞生物のクラミドモナスや、ボルボックスよりもさらに細胞数の少ない群体性の藻類も知られている。このような単細胞生物から多細胞生物への進化(多細胞化)の系譜を辿れるかのようなグループは極めて稀であり、ボルボックスとその仲間は多細胞化を解き明かす鍵といっても過言ではない。私たちは、ゲノム解析や遺伝子解析などの分子生物学的研究を通して、この多細胞化の進化過程を分子レベルで理解しようと試みている。 緑藻ボルボックス(直径0.1〜0.5 mm)は、球面を作る2千個の小さな体細胞と、約20個の大きな生殖細胞からなり、多細胞生物としては最もシンプルな体制をしている。ボルボックスの近縁種には、単細胞生物のクラミドモナスや、ボルボックスよりもさらに細胞数の少ない群体性の藻類も知られている。このような単細胞生物から多細胞生物への進化(多細胞化)の系譜を辿れるかのようなグループは極めて稀であり、ボルボックスとその仲間は多細胞化を解き明かす鍵といっても過言ではない。私たちは、ゲノム解析や遺伝子解析などの分子生物学的研究を通して、この多細胞化の進化過程を分子レベルで理解しようと試みている。【かたち作りの遺伝子と生殖細胞と体細胞の違いを決める遺伝子を鍵に研究を展開】 ボルボックスの生殖細胞は分裂して胚を作る。この胚は成体のボルボックスと異なり、将来の生殖細胞になる予定の大きな細胞を外側に持ち、成体と球面の表裏が逆転している。ボルボックス胚には一カ所穴が開いており、その部分から外側へとめくれ返っていき、最終的に表と裏が逆転し(反転)、成体の形態になる。その後、生殖細胞はより大きく成長し、体細胞は糖タンパク質を分泌する事により、球状のボルボックスのカラダを大きく構築していく。私たちは、ボルボックスの形態形成運動である反転や生殖細胞と体細胞の分化に働く遺伝子を見出だし、細胞内での働きを研究しています。さらに、そのようなボルボックスのカラダを作るのに必須な遺伝子が、単細胞生物の持つ遺伝子からどのようにして進化してきたのかを明らかにしようとしています。 |
発生過程の脳組織では、ニューロンを作る元となる神経前駆細胞が決められた時期に正しい数と種類の細胞を産生し、やがて機能的な脳が作り上げられます。多様でダイナミックな細胞集団から成る脳組織は、どのように細胞たちを統制し、秩序立った組織形成を可能にしているのでしょうか。本研究室では、細胞の形・動き・力に着目し、主にマウスの脳を対象として、スライスカルチャーシステムを用いたタイムラプスイメージングや、子宮内エレクトロポレーション法を用いた遺伝子導入などの技術を駆使しながら、脳組織づくりの原理を探ります。
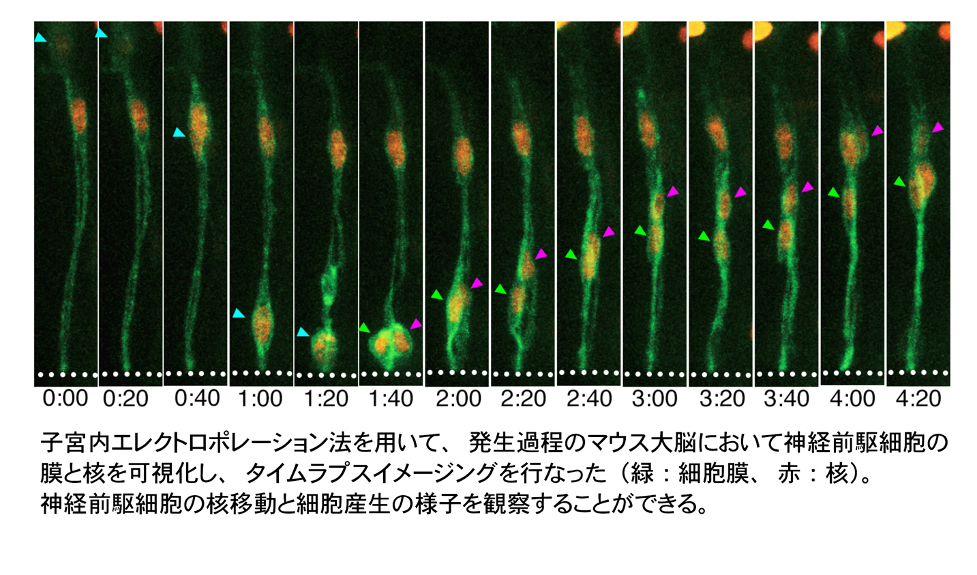
|
| 【眼以外での光感覚に関する研究】 動物は光を様々な場面で利用しています。最も代表的な光の利用法として、眼でものの色や形を見る「視覚」があります。動物は、視覚以外にも、環境の光情報をもとに、時刻や季節などを知ることができます。このような、視覚以外の目的で光を受容することを、「非視覚」の光受容と呼びます。非視覚の光受容のために、哺乳類を除く多くの脊椎動物は、眼以外にも、松果体や脳深部などに光を受容する器官(細胞)を持っています。私たちは、非視覚の光受容、特に、眼以外での光感覚に着目し、その光受容メカニズムやそれらが制御する生理機能について調べることで、動物がどのように光と関わっているのか、その生物学的意義を明らかにしたいと考えています。 〈図〉 下等脊椎動物における眼外の光受容器官 
|
 1. 植物オルガネラの増殖、分化の制御機構と進化
1. 植物オルガネラの増殖、分化の制御機構と進化植物の細胞の中には、光合成を営む葉緑体(色素体)と、呼吸を営むミトコンドリアが存在します。これらはそれぞれ光合成機能および好気呼吸機能をもつバクテリアの細胞内共生によって誕生したと考えられており、それぞれ独自の遺伝子DNAをもちます。植物細胞の増殖/分化の過程で、これらの細胞小器官は遺伝子の複製や遺伝子発現を行いながら半自律的に増殖し、細胞のタイプごとに、また環境変化に応じて、様々な機能を営むように分化しています。私たちの研究室では、これら細胞小器官の増殖/分化がどのように調節されているのか、主に「細胞小器官遺伝子の複製制御」と「細胞小器官遺伝子の転写制御」の二つの観点から研究を進め、そのような制御機構がどのようにして作り上げられてきたのか(進化的背景)、また、植物の生存にとってどのように役立っているのか(適応的意味)、といった問題について考察しています。 2. 化学物質を介した植物個体間相互作用 ―他感作用― 「ある植物が放出した化学物質によって、他の植物が何らかの作用を受ける現象」を「他感作用(アレロパシー)」といいます。自然界では阻害的な作用を与える例、すなわち「ある植物の周囲に他の植物が生えにくい現象」の観察例が多いようです。自分自身の周囲から他の植物を排除することは、植物が生きていくために必要な光や水、無機養分を獲得する上で有利に働くと考えられます。私たちの研究室では「他感作用が実際にはどのように、そしてどの程度、植物の生存に貢献しているのか」という生理/生態的な意義と、「他感作用をもたらす化学物質はどのようなメカニズムで他の植物の生育に影響を及ぼすのか」という生理/生化学的な機構の二つの観点から研究を進めています。 |
|
1. 植物細胞の時空間ダイナミクス制御に関する研究 細胞分裂停止後、殆ど大きさを変えない動物細胞とは対照的に、植物細胞は分裂停止後、100倍以上にも体積を増大させます。この『細胞成長』は、地球上のバイオマス生産の80%以上を占める植物の高い成長能力の主要因です。植物の旺盛な細胞成長は、『巨大液胞』という細胞小器官(オルガネラ)の発達によって駆動されます。その一方で、巨大液胞は細胞体積の90%以上を占めるまで大きくなるため、細胞内の時空間ダイナミクスを妨げる物理的障壁ともなりえます。このような「諸刃の剣」となりうる巨大液胞の動態を、植物細胞がどのように制御し、細胞内構造を最適化しているかはよく分かっていません。私たちは、この疑問の解明に、高度なイメージング技術を中心に取り組んでいます。 2. バイオスティミュラントの開発 化学系研究者との異分野融合研究を通して、植物に環境ストレス耐性を付与する化合物(バイオスティミュラント)の開発に取り組んでいます。モデル植物での開発研究、有用植物での実証実験を経て、社会実装を目指し、植物研究を通じてSDGsに貢献していきたいと考えています。 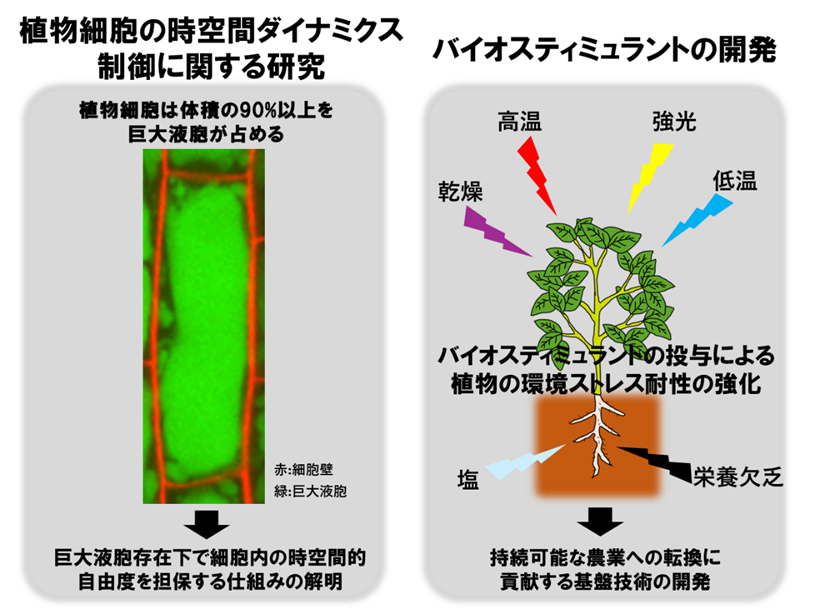
|
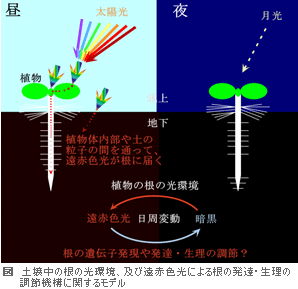 光は植物にとって、光合成のエネルギー源であるとともに、環境を知るための大切な情報源です。地上部(葉や茎など)は光の影響を直接受け、光合成や生長速度を変化させることができます。一方、根の形態形成や生理も光環境に応じて変化します。 光は植物にとって、光合成のエネルギー源であるとともに、環境を知るための大切な情報源です。地上部(葉や茎など)は光の影響を直接受け、光合成や生長速度を変化させることができます。一方、根の形態形成や生理も光環境に応じて変化します。例えば、光は根の細胞伸長を抑制し、根毛形成を促進します。また根による水の吸収と輸送は、地上部での水需要、すなわち光合成や生長(細胞の増殖・伸長等)と調和して行われます。 私達の研究室では、土壌中の根が「地上の光環境の変化」及び「光によって引き起こされた地上部(葉や茎など)の変化」をどのような仕組みで受け取り、根の遺伝子発現や発達・生理の調節をしているのかに着目して研究を行っています。 植物体の地上部に光が照射されると、図のように、一部の光は植物体内を伝わって根まで到達します。この光が遠赤色光の成分に富むことから、土壌中(特に地表面近く)の根は、日中は遠赤色光によって照らされていると考えています。 この遠赤色光の役割を明らかにするために、cDNAマイクロアレイ解析を行い、シロイヌナズナの根の遠赤色光応答遺伝子を特定したところ、6つの水チャネルタンパク質 (アクアポリン:TIP1;1, TIP1;2, TIP2;1, TIP2;2, PIP1;2, PIP2;3) や感染特異的タンパク質(pathogenesis-related protein 1 : PR1)に類似するタンパク質 の遺伝子が含まれていました。 現在は、これらの遺伝子の遠赤色光による発現調節の仕組みや根における機能を明らかにするための研究に取り組んでいます。 また、シロイヌナズナの根毛形成促進突然変異体を用い、光による根毛形成誘導機構を明らかにするための研究も行っています。 |
| 逃げ方の最適化は、動物が持つ根源的な行動です。例えば私たちは、「熱い」という感覚に対して、アイロンの蒸気程度であれば「手を引っ込める行動」ですませますが、灼熱の火事では「全力で逃げる」行動に切り替わります。行動の決定には状況や性差、育った環境など様々な要因が絡み、不具合があると精神・神経疾患にも繋がります。本研究室では、行動を最適化する神経回路がどのように作られ、使われ、適切な行動を導くのかに焦点を当て、全神経のシナプスと遺伝子発現情報が同定されている唯一のモデル生物、線虫 C. elegans を用いて解析します。 研究テーマ


|
 (1)光環境の季節変動に応じた植物の資源利用と繁殖特性 (1)光環境の季節変動に応じた植物の資源利用と繁殖特性落葉広葉樹林では、林冠木の展葉と落葉により林床に届く光は変動します。光資源の季節変化がある中で、林床植物がどのように対処しているのか、繁殖戦略を明らかにするために、植物の生理特性と繁殖特性の関連性を調べています。種子生産の二大制限要因である花粉制限(交配機会による制限)と資源制限(資源量による制限)を結びつけることで、光環境の季節変動が林床植物の生理・繁殖特性に強い選択圧として作用している実態が明らかになってきました。さらに、植物の生理的な特徴を捉えることで、選択圧に対する植物の反応は、個々の植物種の生活史特性(葉寿命や繁殖スケジュール)を反映して多様であることを示しました。 (2)花形質を介した植物と昆虫の生物間相互作用 植物は自ら動き回ることができないので、他個体との交配を実現するには、花粉媒介者を必要とします。植物にとって多様な花形質は、その花粉媒介者を誘う戦略です。その花形質の一つで、ユニークな形質として花色変化があります。意外と多くの植物で見られるこの開花中に花色を変える性質は、花粉媒介動物の行動を制御して花粉輸送効率を高める戦略として機能していることを、野外観察と操作実験により明らかにしました。さらに、花色変化が様々な分類群で見られつつも、植物界では一般的な形質ではないことに着目し、フィールド調査研究に基づくコンピューターシミュレーションによりこの形質の進化条件を示した研究も行っています。 (3)繁殖特性が種間相互作用を介して群集や生態系にどのように作用するのか 植物の繁殖プロセスを介した個別の生物間相互作用が、群集レベルへどのように波及するのか(カスケード効果)を明らかにすることを目的としています。例えば、多くのポリネーターはジェネラリストであり、複数の植物種を同時に利用します。従って、ある時期の花資源量がポリネーターの個体数や分布に影響することにより、別の時期の植物への送粉サービスに影響する可能性が十分考えられます。つまり、同じポリネーターを共有する植物種間には、間接的な相互作用が存在することになります。従来の進化生物学では、ある花形質や防御形質の機能がその機能を有する植物の適応度にどう作用するかが中心課題でした。生態系を物質循環やエネルギーの流れに着目するだけでなく、生態系ネットワークが駆動する動的な系として捉えることで、進化生態学と群集生態学を繋ぎ、生物の形質進化と生物多様性の維持創出機構の統一的な解明を目指しています。 |
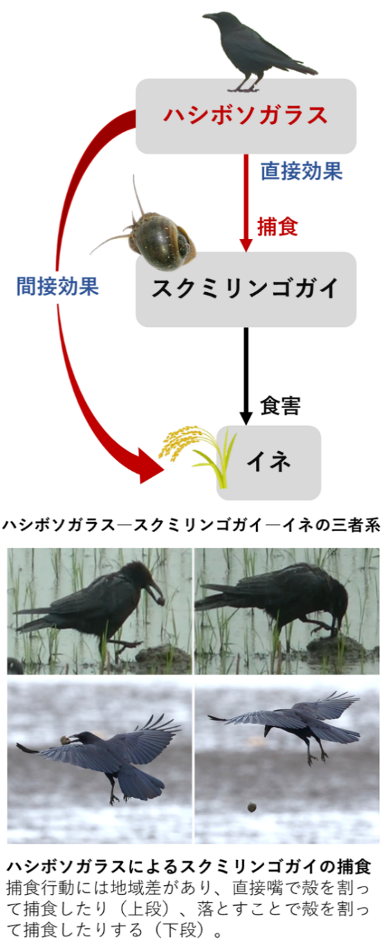 ハシボソガラスースクミリンゴガイーイネの三者系を用いて、動物の学習や適応行動について生態学的および心理学的な観点から研究を行っています。野外においてカラスがどのように学習を行い、その学習が他の個体に伝播するのか、さらにその結果が生態系全体に与える波及効果について調査しています。
ハシボソガラスースクミリンゴガイーイネの三者系を用いて、動物の学習や適応行動について生態学的および心理学的な観点から研究を行っています。野外においてカラスがどのように学習を行い、その学習が他の個体に伝播するのか、さらにその結果が生態系全体に与える波及効果について調査しています。* 鳥類における適応行動、学習や他個体への行動伝播 カラスによるスクミリンゴガイの捕食行動は、特定の地域でしか観察されていません。捕食のある地域の中でも地域差があり、直接嘴で殻を割って捕食したり、落とすことで殻を割って捕食したりします。落とす際には、落下地点の基盤の硬さに応じて落とす高さを適応的に調節することが分かってきました。これらから、カラスによる貝捕食には学習が関与しており、貝捕食の様式が特定の地域の群れで社会的に伝播しているのではないかと考えています。 鳥類のなかでも学習能力に秀でたカラスが、野外でどのように学習を成立させ、その学習が他の個体に伝播していくのかを生態学的および心理学的な観点から解明することを目指しています。 * 学習が生態系全体に与える波及効果 カラスがスクミリンゴガイを捕食する一部の地域では、カラスの捕食率が2日間で80%以上に達することが分かりました。これは、カラスの捕食が直接的にスクミリンゴガイの個体数を低減させることを示唆しています。また、カラスの捕食がスクミリンゴガイの行動を変化させイネの食害を減少させる間接的な効果もあることが分かってきました。このように、カラスによる捕食の学習が、外来種侵入のような現代の生態系に与える波及効果についての理解を深めることを目指しています。 スクミリンゴガイは、国際自然保護連合(IUCN)の「世界侵略的外来種ワースト100」にも含まれる有害な外来種であり、その防除が重要な課題となっています。得られた知見をもとに、外来種管理におけるカラスの活用についても取り組んでいきたいと考えています。 |
 (1)陸水生態系の生物多様性維持機構に関する研究
(1)陸水生態系の生物多様性維持機構に関する研究陸水生態系の生産者である藻類、藻類食者、捕食者の被食捕食関係や摂食行動、分布動態等に着目し、食物網内での相互作用と生物多様性維持機構との関係について研究を行っています。実証的手法と同時にメタデータ解析も行っており、最近の研究では,陸水生態系では捕食者の種多様性が減少すると被食者の種多様性も大きく減少することを明らかにしています。 (2)ダム河川における課題抽出や環境修復策に関する研究 日本では、流程にダムをもたない河川はほぼありません。ダムは河川という流水域に止水域を創出することで河川生態系に大きな影響を与えています。私は主に底生動物を対象として、河川の土砂レジームの改変が食物網や生物多様性にどう改変しているのか調べています。支川流入によるダム影響の緩和や、土砂還元など環境修復策の効果についても検証しています。 (3)里地の小さな陸水域における生物多様性保全に関する研究 ため池は、里地の生物多様性を補償する陸水域として重要性が認識されてきています。また、山裾や農地からの滲み出しによる小さな湿地には希少種が多く生息するため、保全が急務となっています。私はため池の斜面方位や池内のハビタット多様性による補償効果について研究を進めています。また、近年開発されつつある環境DNA技術を、湿地やため池等の野外に適用する研究にも取り組んでいます。 |
 有性生殖が生物の分布と数にどのように影響するのかに興味をもって研究をしています。大きく分けると以下の3つの研究テーマに取り組んでいます。
有性生殖が生物の分布と数にどのように影響するのかに興味をもって研究をしています。大きく分けると以下の3つの研究テーマに取り組んでいます。
1.繁殖形質の進化 繁殖形質はふつう、種内のオスとメスの相互作用によって進化します。どのような繁殖形質が進化してくるのかを理解するために、性選択や性的対立の研究を行っています。最近はおもに、花寿命をめぐる種子親と花粉親の利害対立について、カンサイタンポポやシロイヌナズナを用いて研究しています。特に、花粉が受粉相手の花を無理やり閉じさせたり枯らせたりしているのではないか、という仮説に基づいて実験を進めています。 2.繁殖干渉の行動メカニズム 非常に近縁な2種はふつう異所的に分布していますが、何らかの理由で二次的接触が起きることがあります(例:人為的な移入など)。近縁種が二次的に接触すると、種間交尾や種間送粉などが起き、これが個体の適応度を下げることがあります。このような現象を繁殖干渉と呼びます。繁殖干渉が、繁殖形質とどのように関係しているのかなどに興味をもって研究しています。過去には昆虫のマメゾウムシ類を用いて実験をしていました。 3.繁殖干渉の進化生態学的な帰結 繁殖干渉がどういった生態学的な帰結をもたらすのかにも興味をもっています。繁殖干渉は個体群動態に影響を与えたり、繁殖干渉を避けるような適応を進化させる選択圧になると考えられます。たとえば、繁殖干渉を避ける適応としてどのような形質が進化しやすいのかについて数理モデリングを用いた研究を行なったりしています。こうした理論予測の実証的な検証にも取り組んでいきたいと考えています。 |
 1.侵入種スクミリンゴガイの生態 1.侵入種スクミリンゴガイの生態南米原産の淡水巻貝スクミリンゴガイは世界的な稲の有害動物で、世界と日本の侵略的外来種ワースト100の両方にリストされている唯一の淡水無脊椎動物です。この貝の生態を解明し、防除法の開発に役立てる研究を行っています。近年の主なテーマは以下の通りです。
フジツボ類には、雌雄同体、雌雄同体+矮雄、雌+矮雄という三つの性表現があり、種ごとにどの性表現をとるのか決まっています(ダーウィンの発見)。なぜこのように多様な性表現をもつのか調べています。 3.エボシガイ類の自然史 フジツボの仲間であるエボシガイ類には、潮間帯〜深海までの岩場・海表に漂う流木・他生物などさまざまな基盤を生活の場として、400種以上が知られています。甲殻類のくせに移動性を捨ててしまった彼らがどのような生活を送っているのか調べています。 4.雌雄同体動物の繁殖生態 アメフラシなど雌雄同体動物の繁殖を調べています。なぜ雌雄同体になるのか、雌雄同体特有の繁殖事情(オスとしてふるまう? メスとしてふるまう?)、どのように配偶相手を選択するか、などを研究しています。 |








