最新の研究紹介
数理の視点から生命現象を理解する試み
 [ リリース: 2019.02 ]
[ リリース: 2019.02 ]
奈良女子大学 化学生物環境学科 環境科学コース 教授 高須夫悟
http://gi.ics.nara-wu.ac.jp/~takasu
数理的手法を用いて様々な生命現象を研究する数理生物学を専門としています。特に、生き物の数の時間変化(専門用語で個体群動態と呼ぶ)に注目した数理的研究をしています。数理的手法とは、数式やアルゴリズムと言った抽象的なものの見方を通して研究対象をモデル化して解析する手法です。生命現象と言えば一般的には生物学が扱う研究対象になりますが、数理的手法と生物学がどのように結びつくのか、直ぐには想像できない方が少なくはないのでは無いかと思います。ここでは、数理的手法が如何にして生物学と結びつくのかを説明するとともに、私が取り組んできた研究事例をいくつか紹介し、数理的手法と生物学の深い関わりについて知って頂こうと思います。環境科学は人間を取り巻く生物学的環境(生態系)や物理学的環境に関わる研究分野です。環境科学においても数理的手法が役に立つことを知って頂ければと思います。
生物個体が集まったものを生物集団と呼びます。ある生物集団の個体数(集団サイズとも言う)は以下の式に従って変化(増減)することは直感的にも自明でしょう。
[集団サイズの正味の変化] = [出生数] - [死亡数] + [外部からの移入] - [外部への移出]
この式は、物理学における質量保存の法則と同じく、注目する生物集団の個体群動態を「厳密に」記述します。
全ての生物個体は、無性生殖・有性生殖、動物、植物など繁殖様式の違いはあるものの一定の規則に従い子孫を残し(出生)、寿命が尽きます(死亡)。動物のように空間を移動できる生き物もあります。この式を用いると、出生と死亡(そして移動)がどのように起こるのかがわかれば、これを基にして集団サイズの正味の変化が計算でき、集団サイズの時間変化(個体群動態)が理解できることになります。
個体群動態の最も身近な例として地球人口(地球上の人間の数)が挙げられます。地球人口は2018年に76億を超えたと推定されており、今後も増加し続けることが確実とされています。地球人口が増え続けているのは、毎年の出生数が死亡数を上回っているからに他なりません(地球内外への移動は無視できる)。一方で、日本の人口は過去10年の間減少し続けており、この傾向は当面続くと予想されています。ある年の出生数は出生率(女性一人が産む赤ちゃんの数)と女性人口の積、死亡数は死亡率(一人が病気や事故などで死亡する確率)と人口の積として与えられ、これを元に人口の正味の変化(プラスかマイナス)が計算出来ます。出生率と死亡率は年齢に依存します。具体的には、各年齢の人口をベクトルで表し、年齢に依存する出生率と死亡率を要素に持つ行列を用いて翌年の人口ベクトルを計算するモデルとなります(線形代数の知識が必要)。
ここまでは生物個体を単位として個体数の変化に注目してきましたが、「細胞」を注目する単位とすれば、上式を用いて、体内の特定の細胞(例えばがん細胞)の動態を数理的に記述できることになります。また、麻疹やインフルエンザといった感染症のように、生物個体が様々な状態(未感染で今後感染しうる感受性状態、感染状態、快復後の免疫獲得状態など)を持つと考えると、感受性個体は感染者と接触することで感染状態となり(感染症の出生)、感染者の快復(感染症の死亡)を用いて、感染者の個体群動態としての感染症の流行が数理的に記述できることになります。
個体群動態の数理で取り扱う最も単純なモデルは、微分方程式(連続時間の場合)や差分式(離散時間の場合)を用いて記述されます(解析学の知識が必要)。注目する集団が年齢(何歳の個体がどれだけいるか?)や空間分布(どこにどれだけいるか?)、そして感染症のように様々な属性を持つ場合(このような集団を構造を持った集団と呼びます)は、行列モデルや偏微分方程式、積分方程式など、さまざまな数式を用いて個体群動態を記述することになります。対象となる個体群動態を一旦「数理モデル」として記述してしまえば、後は数学の知識を用いてこのモデルの振る舞いを解析することになります。モデルが複雑な場合は、計算機を用いて数値的もしくやシミュレーションによって振る舞いを解析することになります(プログラミングの知識が必要)。また、細胞や個体の出生死亡といった短い時間スケールではなく、突然変異の出現といったより長い時間スケールに注目すると、ある形質の進化に関する進化動態を扱うことになります。
数理生物学で用いる数理的手法は、皆さんがおなじみの物理学(ニュートン力学、電磁気学、統計力学、量子力学など)や化学(反応速度論など)の分野では必須の手法になっていますが、生命現象にも同様に応用可能かつ有用であることがお分かりになって頂けたかと思います。世間一般では、生物学(特に高校までの生物学)は数理的手法の対局にあるようにとらえられている節があるのですが、決してそうではないのです。
私はこれまで数理的手法を用いて様々な生物学の問題に取り組んできました。ここでいくつかの事例を紹介したいと思います。
まず最初に、鳥類育児寄生の進化動態に関する数理的研究が挙げられます。育児寄生は択卵ともよばれる繁殖形態であり、他個体の育児労働に寄生する繁殖方法です。択卵を行う鳥としてはカッコウ Cuculus canorus が有名です。カッコウの択卵を受け入れた宿主が自分とは似ても似つかず大きく育ったカッコウ雛に餌をやる写真は、自然界の不思議の一つとして広く知られています。しかし、鳥学者の研究によると、カッコウの択卵を認識して排除する能力を持つ宿主の存在が知られています。カッコウの択卵を受け入れると宿主は自分の子孫を残せないため、自然選択により、宿主はカッコウ卵と自分の卵の色や模様の違いを手がかりにして択卵を認識排除する択卵対抗手段を進化的に獲得すると考えられます。また、宿主の択卵対抗手段に対抗して、卵の色や模様が宿主卵にそっくりな卵擬態をもつカッコウの存在が知られています。宿主と択卵鳥が互いに対抗手段を進化させる過程は共進化と呼ばれ、共進化動態の結末がどうなるかに関しては様々な数理的研究が行われています。この共進化動態に関して鳥学者と共同して一連の数理的研究を行い、択卵対抗手段の宿主集団中での進化、卵擬態の進化、そして、宿主と寄生者集団中の卵模様の多型維持に関する数理的研究を行いました(Takasu 2003, Takasu et al. 2009, Fossøy et al., 2016, Liang et al. 2017など)。
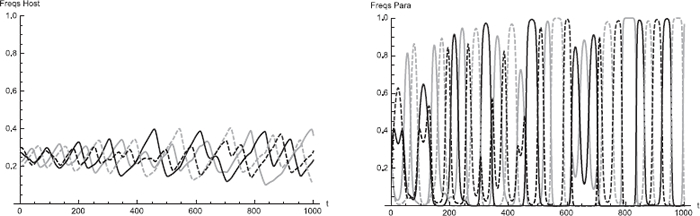
図1: 宿主と寄生者集団中の卵多型の頻度動態の一例(Liang et al. 2017より抜粋)。両集団中で4つの卵形質頻度(背景色が白・薄青、斑点が有り・無しの4通り)が振動して共存する(左が宿主、右が寄生者)。縦軸は頻度、横軸は時間(世代)。
次に、樹木マツの病気の「マツ枯れ」(マツ材線虫病)に関する数理的研究が挙げられます。マツ枯れはマツノザイセンチュウというセンチュウがマツに感染することによって引き起こされます。センチュウはそれ自身では感染マツから感受性マツに移動できないのですが、マツノマダラカミキリによって媒介されることが明らかになっています。健全な松は松脂のためカミキリムシの産卵には適さないのですが、センチュウが感染して衰弱したマツは松脂の滲出が停止し、カミキリムシの格好の産卵場所となります。つまり、センチュウはカミキリムシに新たな産卵場所を提供し、カミキリムシはセンチュウを健全マツに媒介するという、相利共生の強固な関係が形成され、このことがマツ枯れの急速な拡大につながったと考えられています。マツ枯れは、おそらく感染したマツ材の貿易により、現在では韓国や中国といったアジア地域、そしてヨーロッパにまで拡大しており、いまだ深刻な被害をもたらしつつあります。マツ枯れの拡大に関して、単純な差分式を用いた研究を皮切りに、カミキリムシの空間移動を考慮した研究へと、マツガレがどのように空間的に拡がるかに関する一連の数理的研究を行いました(Yoshimura et al. 1999, Takasu et al. 2000, Takasu 2009)。

図2: マツガレの空間的拡大シミュレーションの一例(Takasu 2009より抜粋)。左上から右下へそれぞれ t = 0, 10, 20, 30, 40, 50年後の枯死マツ・感染マツの空間分布。灰色が感染後に枯死したマツ、黒が当年に感染したマツ。
この他にも、日本では特定の高山帯のみに生息する絶滅危惧種のライチョウ Lagopus muta の存続可能性分析を行った数理的研究があります(Suzuki et al. 2013)。日本のライチョウは現在ではごく少数の高山帯のみに生息し、総個体数は2000羽を下回っているとされています。存続可能性分析とは、希少種など個体数が少なくなった生物集団の絶滅リスク評価を行う解析手法です。乗鞍岳のライチョウ集団に関しては、ライチョウ個体の年間生存確率、雌の一服卵数(一度に産む卵の数)、各卵の孵化確率、巣立ち確率などの個体群パラーメータが長年の野外調査により推定されており、この値を用いたシミュレーションを行うことで乗鞍集団が将来絶滅する確率などを計算することが出来ます。本研究で想定した個体群パラメータの下では、乗鞍岳集団は安定しており30年後も存続する可能性が高いことを示しました。しかし現実の系では各高山帯に生息するライチョウはある頻度で移動分散することが示唆されています。複数集団間の移動分散が系全体の存続可能性に与える影響は無視することが出来ず、今後この効果を考慮した研究を進める必要があります。
生物学はミクロからマクロスケールにいたる様々な生命現象に関して数理的研究を行う上で研究事例の宝庫とされています。数理的手法を用いた研究が今後ますます注目され、興味深い現象の解明につながることが期待されています。
参考文献
1. Wei Liang, Canchao Yang, and Fugo Takasu (2017) How can distinct egg polymorphism be maintained in the rufescent prinia (Prinia rufescens)-plaintive cuckoo (Cacomantis merulinus) interactions- a modeling approach. Ecology and Evolution 1-8. DOI: 10.1002/ece3.3090.
(https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5551090/)
2. Frode Fossøy, Michael D. Sorenson, Wei Liang, Torbjørn Ekrem, Arne Moksnes, Anders P. Møller, Jarkko Rutila, Eivin Røskaft, Fugo Takasu, Canchao Yang and Bård G. Stokke (2016) Ancient origin and maternal inheritance of blue cuckoo eggs. Nature Communications 7, Article number: 10272. doi:10.1038/ncomms10272
(https://www.nature.com/articles/ncomms10272)
3. Ayaka Suzuki, Atsushi Kobayashi, Hiroshi Nakamura, and Fugo Takasu (2013) Population viability analysis of the rock ptarmigan Lagopus muta japonica in Japan. Wildlife Biology 19:339-346.
(https://bioone.org/journals/Wildlife-Biology/volume-19/issue-4/13-021/Population-viability-analysis-of-the-Japanese-rock-ptarmigan-i/10.2981/13-021.full)
4. Takasu F. (2009) Individual-based modeling of the spread of pine wilt disease: vector beetle dispersal and the Allee effect. Population Ecology 51:399-409.
(https://link.springer.com/article/10.1007/s10144-009-0145-5)
5. Takasu F. (2003) Co-evolutionary dynamics of egg appearance in avian brood parasitism. Evolutionary Ecology Research 5: 345-362.
(http://www.evolutionary-ecology.com/issues/v05n03/eear1470.pdf)
6. Takasu F., N. Yamamoto, K. Kawasaki, K. Togasi and N. Shigesada (2000) Modeling the range expansion of an introduced tree disease. Biological Invasions 2(2):141-150.
(https://link.springer.com/content/pdf/10.1023/A:1010048725497.pdf)
7. Yoshimura A., K. Kawasaki, F. Takasu, K. Togashi, K. Futai and N. Shigesada (1999) Modeling the spread of pine wilt disease caused by nematodes with pine sawyers as vector. Ecology, 80(5), pp. 1691-1702.
(https://esajournals.onlinelibrary.wiley.com/doi/full/10.1890/0012-9658%281999%29080%5B1691%3AMTSOPW%5D2.0.CO%3B2)
